What Causes Postharvest Senescence in Fresh Produce

Dr. Vijayalaxmi Kinhal
March 31, 2025 at 3:54 pm | Updated March 31, 2025 at 6:43 pm | 7 min read
- Senescence is one of the top two postharvest processes that cause food loss of fresh produce by curtailing shelf life.
- Ethylene production, programmed cell death, and cellular energy supply are significant processes associated with senescence in postharvest fresh produce.
- The mechanisms by which the three processes cause senescence are not entirely understood.
Premature senescence and ripening are two crucial postharvest physiological processes that lead to quality decline and loss of fresh produce. In 2017, about 15-20% of the postharvest losses were due to senescence. Hence, knowing the process and factors that influence senescence can help delay and improve fresh produce quality and shelf life. This article will cover some essential processes known to control senescence.
Extending Shelf-life
The pre-harvest factors and processes that influence the shelf life of fresh produce have received considerable attention. The same is also true of postharvest ripening, but not senescence.
Postharvest senescence is delayed through chemical and physical treatments and a controlled or modified atmosphere for postharvest storage. However, the treatments and storage conditions can also be stressors and induce changes that deteriorate fresh produce quality. The treatments and storage conditions reduce respiration, ethylene production, aging, and associated metabolic changes to extend shelf life. Still, they can lead to chilling injury and disorders due to high carbon dioxide, high ethylene, low oxygen, and less light during storage.
Subscribe to receive our monthly round-up of articles.
The symptoms of these quality declines are known, like browning, yellowing, necrosis, etc., but not the cellular processes.
Some significant factors connected with senescence are ethylene, programmed cell death, and energy availability.
Ethylene
Ethylene is a significant factor in plant senescence, including the postharvest stages. Ethylene is known to increase respiration, the activity of enzymes such as peroxidase, lipoxidase, phenylalanine ammonialyase, permeability, cell decompartmentalization, and changes in auxin transport. Ethylene acts with another senescence promoter, abscisic acid (ABA). Senscence inhibitors like auxins, cytokinins, and gibberellins can also be involved.
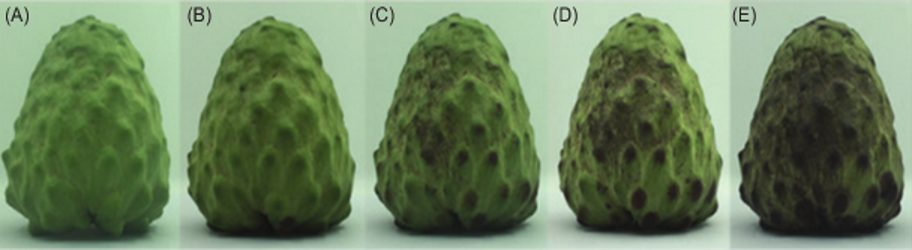
Figure 1: “Five stages of postharvest atemoya fruit senescence. (A) at harvest, (B) 1d, (C) 3d, (D) 5d, (E) 7d,” Gong et al. (2020). (Image credits: DOI: https://doi.org/10.15586/qas.v12i4.786).
Ethylene action accelerates fresh produce deterioration to shorten shelf life. The phytohormone is necessary to ripen mainly climacteric fruits, but all fresh produce is sensitive to its role in senescence. The phytohormone also adversely affects fresh produce quality and physiology and promotes postharvest diseases.
Loss of quality: Ethylene effects on postharvest fresh produce include changes in color, texture, flavor, potato tuber sprouting, etc.
- Color: Ethylene is involved in chlorophyll degradation of vegetables like greens, cabbage, broccoli, cucumber, etc. Even small amounts of 1-5 ppm can cause degreening in some cabbage.
- Texture: Senescence reduces firmness and rind thickness, for example, in watermelons.
- Abscission: Ethylene causes the abscission of leaves in cabbage, foliage plants, flowers, broccoli florets, etc.
- Flavor: Ethylene causes unflavorable flavors, such as bitterness in sweet potatoes, carrots, cabbage, etc. Cabbage exposed to 100 ppm in controlled atmosphere storage can taste bitter.
- Sprouting: Exposure to 2 ppm ethylene causes tubers to grow in storage, cutting the shelf life of potatoes meant for the table.
Physiological disorders: Ethylene causes several physiological disorders, whose intensity and incidence depend on the commodity’s physiological age, temperature, ethylene concentration, and exposure duration.
- Cut flowers: Ethylene can result in abscission, fading, wilting, and or failure to reopen (sleepiness) of cut flowers.
- Discoloration: Ethylene levels of 50 ppb in a controlled atmosphere can cancel the benefits of cold storage, for example, in kiwifruits, by leading to pulp softening and skin browning. The gas also causes brown stains on lettuce or scalding in apples stored for up to seven months.
Depending on the pathogens, ethylene can promote or inhibit disease development in postharvest fruits. The gas increases fungal growth and increases Botyris cinerea damage in strawberries. However, in some cases, ethylene increases resistance to pathogens, such as sweet potato resistance against Ceratocystis, and inhibits apple rot by Gloeosporiumm album.
Practical Implications
The effects of ethylene senescence on postharvest fresh produce are well-known, even though the mechanisms involved are not. Controlled atmosphere storage and modified atmosphere packaging aim to reduce ethylene levels using scrubbers, scavengers, etc. Factors like warmer temperatures or machinery that could increase the gas levels in a room are also strictly controlled. In climacteric fruits, delaying the respiration rate peak associated with a spurt in ethylene production can extend shelf life.
Programmed Cell Death
In fresh produce that is deteriorating, cells die and disappear. Fluids from the cells leak and fill intercellular spaces to form lesions. These observations indicate that programmed cell death (PCD) is involved during senescence.
PCD is a process strictly controlled by genetics as a response to stress. In the postharvest stage, storage conditions (extreme temperatures, low oxygen, high carbon dioxide), pathogens, mechanical damage, and physical and chemical treatments are stresses that trigger PCD.
There is a dispute about whether senescence and PCD are the same or separate processes, where PCD occurs after senescence. Earlier, the former theory was favored, but many scientists now consider that the phases are not isolated and symptoms of PCD occur during senescence. These symptoms are the expression of genes associated with senescence and PCD, hormonal changes, lytic enzyme accumulation in cell compartments and storage in vacuoles until use during final dismantling, and signaling involving protein and lipid-derived factors.
Several senescence processes are also standard to PCD, like nuclear DNA and chlorophyll breakdown, activation of destruction pathways, cytoskeleton changes, increase in tonoplast and plasma membrane permeability, proteolysis, electrolyte leakage, etc.
Senescence can be considered to be a specific form of PCD.
PCDs in senescence can occur in autophagic processes or vacuolar PCD and apoptosis-like (APL) PCD. See Figure 2 for a schematic depiction of the differences between the two pathways.
- PCD causes the decline of fresh produce quality because it changes physiology and biochemistry, making the product susceptible to pathogens.
- PCD causes browning of the litchi pericarp due to energy deficiency.
- PCD causes physiological deterioration in cassava due to oxidative and protease upregulation processes during cell death.
Practical Implications for the Food Supply Chain
Understanding the role of PCD can help develop optimized or new preservation strategies to control postharvest stress-induced PCD and breeding cultivars with tolerance to storage-induced stress.
A crucial consideration to remember while developing postharvest management is that cell death occurs in three stages. Changes in the initial phases are reversible. The senescence process becomes irreversible at the end of the second stage when the so-called ‘point-of-no-return’ is reached. Steps that can delay the early phases can reverse the development of disorders due to storage stress. For example, intermittent warming can be used during storage to avoid chilling injury due to low temperatures. Or gas composition changes can alleviate the effects of low oxygen and high carbon dioxide.
New postharvest methods must also be tested for any PCD effects. Strategies that target cell death can alleviate the sensitivity to browning that occurs as a result of postharvest stress in whole and cut fresh produce storage.
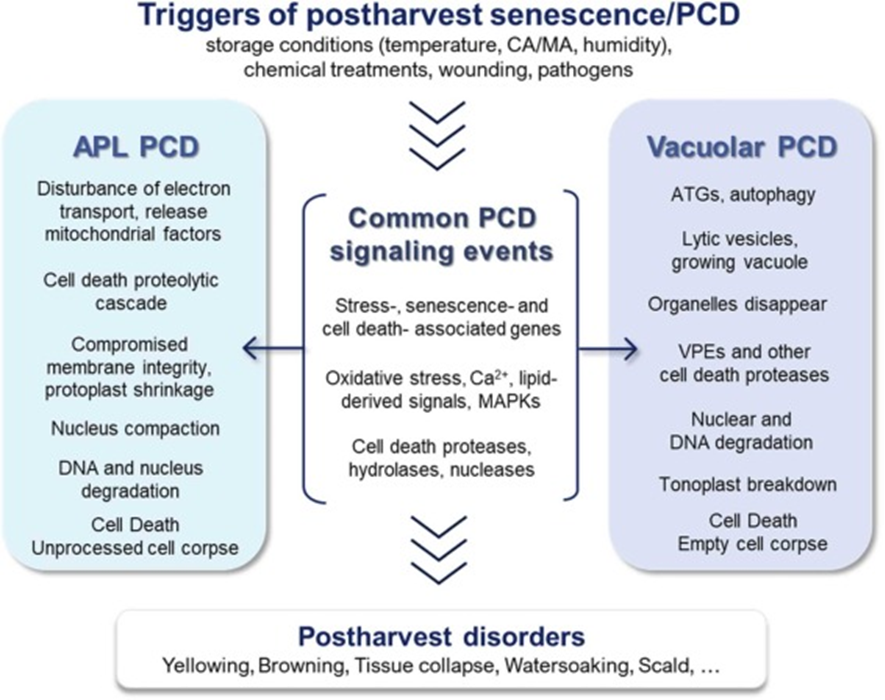
Figure 2.: “Schematic presentation of the processes involved in APL and vacuolar PCD and their common signaling events,” Iakimova et al. 2024. (Image credits: https://www.sciencedirect.com/science/article/pii/S0925521424002552)
Energy
Cellular energy supply is another key factor influencing senescence. Senescence and browning seen in postharvest fresh produce could be due to less or inefficient generation of cellular energy supply. The onset of senescence is associated with low adenosine triphosphate (ATP) levels due to increased membrane permeability and reactive oxygen species (ROS) production. Certain energy levels are necessary for fruits to maintain cell function. Energy status affects the respiratory pathway, ROS production, and integrity of membrane lipids. ROS, membrane damage, and lack of energy can initiate senescence and decompartmentalization of intracellular structures.
However, the required energy levels will differ between climacteric and non-climacteric fresh produce and need further study. In the case of climacteric fruits, the energy levels peak along with respiration and ethylene production before declining.
Evidence for the role of low ATP in senescence has been observed in several species in various storage conditions. For example, ‘Conference’ pear that had high quality after 5-6 months of controlled atmosphere storage had high ATP levels, and their delay in senescence was associated with low demand for energy and increased accumulation of ATP.
However, the changes in ATP levels by storage condition are species-dependent. Litchi fruit exposed to high oxygen increased levels of ATP and adenosine diphosphate (ADP), a precursor to ATP. However, high oxygen reduces banana energy production and accelerates ripening and senescence.
The type of storage matters, too. Modified atmosphere packaging maintains energy levels in cherries better than air storage.
Practical Applications
Applying known information, some recommendations to try to maintain energy levels to delay senescence are discussed below:
- Preharvest: The application of boron and calcium can maintain tissue ATP levels.
- Postharvest: Treatments such as ethylene inhibitors, high oxygen, anaerobic conditions, controlled atmosphere, and modified packaging, providing external carbon and ATP, can improve tissue ATP levels and delay pericarp browning in litchi and longan and internal browning in pears.
- Plant breeding: Senescence in litchi, due to low ATP, was associated with the expression of genes LcAtpB and LcAOX1. So, plant breeding can use them as markers to develop future varieties with more ATP production to delay senescence.
Gas Measurements
Measuring gases in storage and packaging is crucial in all three significant processes controlling senescence. Ethylene must be monitored in ppb and ppm levels for its direct effects. In contrast, oxygen and carbon dioxide must be observed not to cause stress that can damage fresh produce, trigger PCD, or reduce tissue energy supply.
Precision tools that are easy to use, small, and portable are ideal. Felix Instruments Applied Food Science produces such devices that give accurate gas measurements in real time. Some tools only measure ethylene, while others measure all three gases suitable for various supply chain purposes. The fixed gas analyzer for storage rooms can also monitor and regulate temperature and humidity to comprehensively control conditions that cause senescence and reduce food loss.
Learn more about Felix Instruments’ Gas Analysis tools for your supply chain needs.
Source
Chen, L., Wang, M., Wang, H., Zhou, C., Yuan, J., Li, X., & Pan, Y. (2023). Isothermal Storage Delays the Senescence of Post-Harvest Apple Fruit through the Regulation of Antioxidant Activity and Energy Metabolism. Foods, 12(9), 1765. https://doi.org/10.3390/foods12091765
Corpas, F.J., Muñoz-Vargas, M.A., González-Gordo, S. et al. Nitric Oxide (NO) and Hydrogen Sulfide (H2S): New Potential Biotechnological Tools for Postharvest Storage of Horticultural Crops. J Plant Growth Regul 44, 1203–1220 (2025). https://doi.org/10.1007/s00344-023-11150-5
Figueroa, C. R., Jiang, C. Z., Torres, C. A., Fortes, A. M., & Alkan, N. (2021). Editorial: Regulation of Fruit Ripening and Senescence. Frontiers in plant science, 12, 711458. https://doi.org/10.3389/fpls.2021.711458
Gong, X., Wu, X., Qi, N., Li, J., Zhang, H., & Huo, Y. (2020). Changes in the biochemical characteristics and volatile fingerprints of atemoya during postharvest ripening at room temperature. Quality Assurance and Safety of Crops & Foods, 12(4), 26-35. DOI: https://doi.org/10.15586/qas.v12i4.786
Iakimova, E. T., Ty, A. J., Maarten, L. A. T. M., Nicolaï, B. M., & Woltering, E. J. (2024). Programmed cell death and postharvest deterioration of fresh horticultural products. Postharvest Biology and Technology, 214, 113010.
Jiang, Y., Jiang, Y., Qu, H., Duan, X., Luo, Y., & Jiang, W. (2007). Energy aspects in ripening and senescence of harvested horticultural crops. Stewart Postharvest Review, 3(2), 1-5.
Kader, A. A. (1985). Ethylene-induced senescence and physiological disorders in harvested horticultural crops. HortScience, 20(1), 54-57.
Zhu, Y., Du, M., Jiang, X., Huang, M., & Zhao, J. (2022). Nitric Oxide Acts as an Inhibitor of Postharvest Senescence in Horticultural Products. International journal of molecular sciences, 23(19), 11512. https://doi.org/10.3390/ijms231911512
Related Products
Most Popular Articles
- Spectrophotometry in 2023
- NIR Applications in Agriculture – Everything…
- The Importance of Food Quality Testing
- The 5 Most Important Parameters in Produce Quality Control
- Melon Fruit: Quality, Production & Physiology
- Fruit Respiration Impact on Fruit Quality
- Guide to Fresh Fruit Quality Control
- Liquid Spectrophotometry & Food Industry Applications
- Ethylene (C2H4) – Ripening, Crops & Agriculture
- Understanding Chemometrics for NIR Spectroscopy



